![]()


研究室からの風景
マスト細胞の活性化における転写因子XBP1の役割
XBP1は遺伝子発現を制御する転写因子の一種で、機能的タンパク質の生成は、IRE1apha酵素によってXbp1 mRNAのスプライシングが行われることにより制御されています。IRE1alpha-XBP1軸は、小胞体ストレス応答の主経路であると同時に、大量のタンパク質を合成する細胞などの機能や分化にも関わり、B細胞の抗体産生能や樹状細胞の分化など、免疫担当細胞においてもXBP1の寄与が知られています。私たちの研究グループでは、これまでに香料成分から抗アレルギー性物質を探索し、サリチルアルデヒド(SA)がIgE依存的なマスト細胞活性化を抑制することを見出していましたが、その機構は不明でした(Ashikari. T. et al. Int. J. Mol. Sci. 2022)。今回、SAがIRE1alphaのヌクレアーゼ活性を阻害することを根拠に、SAより強い阻害剤3-メチル-6-ブロモ-SA (MBSA)や、IRE1alphaのキナーゼ活性阻害剤KIRA6について調べたところ、MBSA、KIRA6共に、IgE依存的なマスト細胞活性化やマウスのアナフィラキシーを有意に抑制することが明らかとなりました。更に、siRNAを用いてXbp1 mRNAをノックダウンしたところ、ノックダウンの程度と比例して、IgE依存的なマスト細胞の脱顆粒反応が有意に減弱化することが確認され、IRE1alpha-XBP1軸がマスト細胞の機能に関わることが示唆されました。
Kouda, H. et al. Int. J. Mol. Sci. 2026.
論文リンク DOI: 10.3390/ijms27104532
インテグリンCD103が粘膜型マスト細胞や樹状細胞に発現する分子機構
CD103は主に血球系細胞表面に発現するヘテロ2量体構造のインテグリンであり、Eカドへリンと接着する作用によって腸管粘膜への細胞局在を制御します。寄生虫感染への生態防御やアレルギーの発症・病態に関わるマスト細胞のうち、粘膜組織に存在する粘膜型マスト細胞は細胞表面にCD103を発現し、プロテアーゼMCPT1やMCPT2を分泌することを特徴とします。これまでに、私達の研究グループでは、MCPT1/2の遺伝子発現が、転写調節因子GATA2とサイトカインTGF-betaとの協調的な働きによって誘導されることを報告していました(Kasakura, K. et al. J. Immunol. 2020)。一方、今回の研究では、CD103発現がMCPT1/2と同様にTGF-betaによって正に制御されながら、GATA2はむしろ抑制的に作用することが明らかとなりました。更に、その機構を調べた結果、マスト細胞ではCD103を構成するalphaサブユニットの遺伝子発現に必須の転写因子PU.1がGATA2によって抑制されていること、樹状細胞においてもPU.1とTGF-betaがCD103発現を制御していること、などが判明しました。
Ishii, K., Nagata, K. et al. FEBS J. 2025.
論文リンク DOI: 10.1111/febs.70252
IL-33囮受容体の遺伝子発現制御機構
IL-33は、細胞の核内に存在し、細胞損傷などによっても細胞外へと放出されるサイトカインの一種であり、その受容体ST2を発現する顆粒球系のマスト細胞や好塩基球、リンパ球系のTh2やILC2などを活性化し、アレルギー反応や寄生虫感染防御に寄与します。私達の研究グループでは、ST2の発現レベルが高いマスト細胞について、その転写調節機構を報告してきました(Baba, Y. et al. J. Biol. Chem. 2012; Baba, Y. et al. Allergol. Int. 2012)。
今回は、ST2をコードする遺伝子IL1RL1が分泌型の囮受容体となるスプライスバリアントを発現する転写調節機構について調べ、ヒトとマウスのマスト細胞に共通して転写因子GATA2による制御が行われていること、並びにその詳細について報告しました。
Nagata, K., Kasakura, K. et al. Allergol. Int. 2025.
論文リンク DOI: 10.1016/j.alit.2025.04.004
芽胞性乳酸菌の免疫賦活作用
乳酸菌は、乳酸産性能をもつ細菌を指し、その一部は宿主の免疫機能を高めるプロバイオティクスとして用いられます。芽胞を形成する特殊な乳酸産生菌Heyndrickxia coagulans SANK70258は、熱や酸に強いことから食品や飼料への添加利用がなされていますが、宿主の免疫機能に及ぼす影響は不明でした。本研究では、SANK70258菌体が樹状細胞の活性化を発端として、NK細胞のIFN-γ分泌やB細胞のIgA産生を促す高い免疫賦活活性を示すことを見出しました。さらに、SANK70258の菌体構成成分に樹状細胞の受容体を刺激する特徴があることが示唆され、SANK70258を経口摂取したマウスでは腸管内のIgA分泌量が有意に増加することが確認されました。
Ikeda, Y., Yamashita, N., Ito, N. et al. Biosci. Biotechnol. Biochem. 2025
論文リンクDOI: 10.1093/bbb/zbae203
内分泌撹乱物質の受容体を阻害して免疫炎症を抑制するポリフェノールの効果
腸管の樹状細胞は、ビタミンA由来化合物であるレチノイン酸を合成する酵素RALDH2のはたらきにより、炎症や過剰な免疫反応を抑制する制御性T細胞の分化を促します。本研究では、RALDH2の発現亢進活性をもつ食品由来成分の探索を進め、ポリフェノールの一種であるケンフェロールが高い活性を有しており、同じフラボノイド類のケルセチンなどにもケンフェロールに準ずる活性があることを見出しました。その分子機構を解析した結果、ケンフェロールは内分泌撹乱物質の受容体である芳香族炭化水素受容体AhRのアンタゴニストとして機能することや、転写因子PU.1/IRF4を活性化することによって、RALDH2の発現を増強し、Treg誘導促進をもたらすことが判明しました。更に、ケンフェロールやAhRアンタゴニスト化合物が、Treg誘導を介して食物アレルギーを緩和することをマウスモデルを用いて明らかにしました。
Takahashi, M., Nagata, K., Watanuki, Y. et al. Allergy 2025
論文リンクDOI: 10.1111/all.16410
プレスリリース
好塩基球とマスト細胞の分化運命を決定する転写調節機構
好塩基球とマスト細胞は、いずれも造血系幹細胞から発生し、IgE受容体を介したアレルギー反応や寄生虫感染防御応答において重要な役割を果たすといった多くの共通点を持つ一方、生体内局在など明確に異なる性質も有します。本研究では、好塩基球とマスト細胞の相違を生み出す転写調節機構を明らかにするため、好塩基球に特異的なプロテアーゼMcpt8とMcpt11の遺伝子発現制御機構を解析しました。その結果、Mcpt8・11の発現に対し、C/EBPαが転写活性化因子であること、ヒストン脱アセチル化が正に、DNAメチル化が負に寄与するエピジェネティック制御があること、GATA2も転写活性化因子として機能し、TGF-β刺激が誘導因子であること、などが明らかになりました。
Tojima, R., Nagata, K., Ito, N. et al. FEBS Lett. 2025
論文リンクDOI:10.1002/1873-3468.15069
多価不飽和脂肪酸の腸内細菌代謝産物がもつ抗炎症作用
近年、多価不飽和脂肪酸が、摂取した宿主の腸内細菌によって新たな化合物に変換されることが、見出されてきています。本研究では、γ-リノレン酸の二次代謝産物であるγKetoCが、炎症反応を抑制し、炎症性腸疾患の症状を緩和することを報告しています。また、その分子機構として樹状細胞の表面に局在するGタンパク質受容体と、細胞内に存在する転写因子NRF2をγKetoCが活性化する可能性を示すと共に、NRF2欠損マウスではγKetoC摂取による腸炎緩和が抑制されることを明らかにしました。
Ando, M., Nagata, K., Takeshita, R. et al. Front. Immunol. 2024
論文リンク DOI: 10.3389/fimmu.2024.1374425
プレスリリース
短鎖脂肪酸の抗アレルギー作用機構
短鎖脂肪酸は、酪酸やプロピオン酸、吉草酸といった炭素鎖の短い脂肪酸を指し、難消化性食物繊維が腸内細菌によって分解される際に発生する二次代謝産物として腸管内に豊富に存在します。その効果は、腸管に留まらず、全身にわたって免疫恒常性の維持に貢献します。短鎖脂肪酸が抗炎症・抗アレルギー効果を示すことはよく知られていましたが、本研究では、短鎖脂肪酸を摂取したマウスにおいてIgE依存的なアナフィラキシー反応の症状が軽症化することを見出し、その機構解析により、マスト細胞上に発現するGPR109A/HCAR2受容体を介してプロスタグランジン産生が促進され、IgE依存的なマスト細胞活性化が抑えられる仕組みや、短鎖脂肪酸のもつヒストン脱アセチル化酵素阻害活性により細胞表面のIgE受容体量が減少する機序などを明らかにしました。短鎖脂肪酸の抗アレルギー効果が非ステロイド性抗炎症薬(NSAIDs)によって阻害されることも判明し、プロスタグランジン産生を阻害するNSAIDsが一部のアレルギー疾患において禁忌であることにも関わる研究内容となっています。
Nagata, K., Ando, D. et al. J. Immunol. 2024
論文リンク DOI: 10.4049/jimmunol.2300188.
プレスリリース
ポリフェノールの抗アレルギー作用機序
ポリフェノールは植物が産生する二次代謝産物の一種で、摂取したヒトや動物の健康に様々な有益な作用をもたらすフィトケミカルです。ケンフェロールやケルセチンといったフラボノール類が抗アレルギー効果を発揮することは多くの研究から知られていましたが、今回、その分子機構の解析により、ケンフェロールがマスト細胞上のIgE受容体発現レベルを低下させ、細胞内シグナル伝達を抑制する脱リン酸化酵素の発現を増強することによって、IgE依存的なアレルギー反応を抑制することが判明しました。
Nagata, K., Araumi, S. et al. Int. J. Mol. Sci. 2023
論文リンク DOI: 10.3390/ijms24065997.
薔薇の香り成分の抗炎症効果
薔薇の香気成分であるβ-ダマスコンに、免疫細胞による炎症反応を抑制する作用があることが見出されました。その作用機序として、β-ダマスコンが抗原提示細胞である樹状細胞に対して抗酸化反応を誘導するNRF2経路を活性化することが判明し、NRF2欠損マウスの解析から、β-ダマスコンを摂取することによりNRF2依存的に皮膚炎症が抑制されることが示唆されました。
Kodama, N., Okada, H. et al. Front. Nutr. 2023
論文リンクDOI: 10.3389/fnut.2023.1081263.
プレスリリース
抗アレルギー作用をもつ香料化合物の発見
白血球の一種であるマスト細胞は、IgE抗体を介して花粉などのアレルギー原因物質を認識すると活性化し、くしゃみや痒みなどの症状を引き起こす化学伝達物質を放出します。このようなマスト細胞活性化を抑制する物質を探索した結果、植物精油中に含まれ香料として利用されている化合物の一つにマスト細胞の脱顆粒反応やサイトカイン産生抑制作用があり、その投与によりマウスのアナフィラキシー反応が有意に抑制されることが明らかになりました。
Ashikari, T. et al. Int. J. Mol. Sci. 2022
論文リンク DOI: 10.3390/ijms23159926
ケモカインTARCの発現制御機構解析とsiRNA核酸医薬のアレルギー性疾患緩和効果
血球系細胞を誘因する作用を持つケモカインの一つであるTARCは、アトピー性皮膚炎の重症度の診断指標ともなっており、アレルギー疾患との関わりが深い分子です。表皮角化細胞や樹状細胞など、特定の細胞がTARCを分泌しますが、その仕組みはほとんどわかっていませんでした。今回、TARCをコードするCCL17遺伝子を詳しく調べ、細胞によって遺伝子上の異なる領域が活性化に関わっていること、樹状細胞では転写因子PU.1とIRF4による協調的な制御が行われていること、PU.1 siRNAを投与したマウスではアレルギー性喘息が緩和することなどを発見しました。
Ito, N., Sakata, F. et al. Allergy 2022
論文リンク DOI: 10.1111/all.15184
プレスリリース
オピオイドによる免疫応答制御と炎症性腸疾患緩和
感覚や感情に影響を及ぼすオピオイドは、主に中枢神経系に作用すると考えられてきましたが、近年では、免疫細胞を含め、末梢の組織にも受容体の発現が確認されています。今回、オピオイド化合物の一種が、マウスの炎症性腸疾患を緩和すること、その仕組みとして中枢神経を介さずに末梢のマクロファージの炎症反応を抑制し、制御性T細胞分化を促進する可能性があることを見出しました。
Nagata, K. et al. Front. Immunol. 2021
論文リンク DOI: 10.3389/fimmu.2021.730706
プレスリリース
PD-L2の発現制御機構
チェックポイント分子PD-1のリガンドの一つ、PD-L2について、その遺伝子発現制御機構として転写因子同士による協調的な転写活性化の仕組みがあることやエピジェネティック制御が行われていることを明らかにしました。
Inaba, K. et al. J. Immunol. 2020
論文リンク DOI: 10.4049/jimmunol.1901008
プレスリリース
マスト細胞・好塩基球の特異的遺伝子発現
マスト細胞と好塩基球は、IgE抗体に対する受容体(FcεRI)を発現しており、アレルギー原因物質(アレルゲン)の結合により活性化し、アレルギー反応を引き起こします。
このような特定の白血球でのみ遺伝子発現が引き起こされる仕組みとして、下図に示すように造血系細胞に特異的な転写調節因子同士の協調的作用や(Inage, E. J. Immunol. 2014)、抑制作用を持つ共役因子の働きなど(Maeda, K. Blood 2006)を見出してきました。
マスト細胞は、IgE抗体以外にも幹細胞増殖因子(SCF)やIL-33、アナフィラトキシンやATPなど様々な分子に対し、受容体を介して応答し活性化します。私たちはこれら受容体や関連分子の特異的遺伝子発現機構を解析すると共に、アレルギー反応制御法の探索を行っています(Baba, Y. J. Biol. Chem. 2012; Maeda, K. J. Immunol. 2010; Oda, Y. Sci. Rep. 2018)。

樹状細胞の遺伝子発現調節
樹状細胞は、病原体侵入に素早く応答してサイトカインやケモカインを産生することにより自然免疫に関わると同時に、代表的な抗原提示細胞として侵入した異物の情報をT細胞に提示しエフェクター化を促すことによって獲得免疫の方向性を制御する役割を担っています。
私たちはこれまで、樹状細胞の抗原提示機能を司るMHC class II分子や、代表的な共刺激分子であるCD80、CD86の発現制御機構を解析してきました(Kitamura, N. J. Allergy Clin. Immunol. 2012; Kanada, S. Blood 2011)。図に示す通り、転写調節因子であるPU.1が共役因子CIITAの発現を介して間接的にMHC class IIの発現を規定していることや、CD80、CD86遺伝子に対する転写活性化機構の詳細を明らかにしています。PU.1はTh2やTfhの分化誘導に関わる共刺激分子OX40Lの遺伝子発現にも関与していることや(Yashiro, T. Sci. Rep. 2016)、パートナー分子IRF4の遺伝子にも作用して樹状細胞マーカー分子でもあるCD11cの発現を引き起こしていること(Yashiro, T. Int. Immunol. 2017)、核内受容体型転写因子NR4A3の発現を介して樹状細胞の分化や活性化に関わっていることなどを見出しており(Nagaoka, M. J. Immunol. 2017)、引き続き樹状細胞の機能制御に重要な分子について発現制御機構を解析しています。
さらに、自己免疫疾患や移植時の拒絶反応に関わる樹状細胞機能を制御する技術開発に向け、学内外の共同研究も行っています。
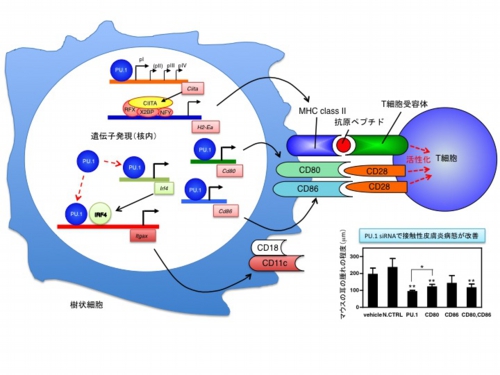
MHC分子発現制御
T細胞への抗原提示はMHC分子を介して抗原由来ペプチドがT細胞受容体へ提示されることによって行われます。全ての有核細胞が発現し得るMHC class Iとは対照的にMHC class II発現はプロフェッショナル抗原提示細胞を中心に、特定の細胞に限定されます。MHC class IIの発現には転写共役因子CIITAの働きが必須であり、CIITAの遺伝子発現は少なくとも3つのプロモーターが細胞種や活性化状態によって使い分けられています。以下のように、これらのプロモーターの制御機構について解析を行っています。