僸僩儗僩儘僂僀儖僗姶愼徢儌僨儖丗僄僀僘儌僨儖
俬丏攚宨
丂儗僩儘僂僀儖僗偑摦暔偵娻傪婲偙偡偙偲偼埲慜偐傜抦傜傟偰偄偨偑丄僸僩俿嵶朎敀寣昦僂僀儖僗乮HTLV乯偼僸僩偵娻乮惉恖俿嵶朎敀寣昦:ATL乯傪堷偒婲偙偡儗僩儘僂僀儖僗偲偟偰弶傔偰摨掕偝傟偨乮侾乯丅偦偺屻HIV偑僄僀僘偺尨場僂僀儖僗偱偁傞偙偲偑柧傜偐偵偝傟丄儗僩儘僂僀儖僗偼僸僩偺昦場僂僀儖僗偲偟偰傕廳梫偱偁傞偙偲偑帵偝傟偨乮俀乯丅偙偺懠偵傕帺屓柶塽昦傗恄宱徢偵儗僩儘僂僀儖僗偺娭梌偑媈傢傟偰偄傞傕偺偑偁傞偑丄尰嵼偺偲偙傠偼偭偒傝偟側偄丅
丂HIV偲HTLV偵嫟捠偟偰偄傞偙偲偼丄廻庡堟偑嫹偔僒儖偺懠偵偼傛偄姶愼宯偑側偄偙偲偱偁傞丅偙偺偙偲偼丄儚僋僠儞傗帯椕栻傪奐敪偡傞偆偊偱戝偒側忈奞偲側偭偰偄傞丅偦偙偱変乆偼偙傟傜偺僂僀儖僗堚揱巕傪摫擖偟偨僩儔儞僗僕僃僯僢僋乮Tg乯儅僂僗傪嶌惢偡傞偙偲偵傛傝廻庡堟僶儕儎乕傪墇偊丄昦懺儌僨儖傪嶌傞偙偲傪帋傒偨丅
丂儗僩儘僂僀儖僗偵傛傞姶愼徢偺儌僨儖偲偟偰Tg儅僂僗傪巊偆偙偲偵偼棙揰偲摨帪偵尷奅偑偁傞偙偲傪巜揈偟偰偍偔昁梫偑偁傞丅棙揰偲偟偰偼丄僒儖傪巊偆応崌偵妑傋丄堚揱妛揑丄旝惗暔妛揑惂屼偑梕堈偱偁傞偙偲丄摦暔偺庢傝埖偄偑梕堈偱偁傞偙偲丄宱嵪揑偱偁傞偙偲丄摦暔垽岇忋偺栤戣偑傛傝彮側偄偙偲側偳偑偁偘傜傟傞丅偟偐傕儗僩儘僂僀儖僗偼帺慠姶愼偺応崌偵傕僂僀儖僗堚揱巕偼廻庡DNA偵慻傒崬傑傟偨忬懺偱堚揱巕敪尰傪峴偆偙偲偐傜丄堚揱巕敪尰偺挷愡婡峔偑帺慠姶愼帪偵嬤偄偙偲偑摿挜偱偁傞丅堦曽丄Tg儌僨儖偼悅捈姶愼條幃傪偲傞偙偲偐傜丄捠忢偺悈暯姶愼偺応崌偲偼堎側傞憻婍偵忈奞偑敪惗偟偨傝丄僂僀儖僗偵懳偡傞柶塽墳摎偑尒傜傟側偄応崌偑偁傞丅傑偨丄僂僀儖僗儗僙僾僞乕偑側偄偨傔偵嵞姶愼偑婲偙傜偢丄姶愼偺奼戝傪尒傞偙偲偑偱偒側偄側偳偺栤戣揰偑偁傞丅廬偭偰丄Tg儅僂僗傪幘姵儌僨儖偲偟偰梡偄傞応崌偵偼丄幚嵺偺昦婥偲斾妑偟偰昦懺偺堎摨傪傛偔嬦枴偟丄儌僨儖偲偟偰偺桳梡惈傪専摙偡傞偙偲偑廳梫偱偁傞丅
丂変乆偼丄HIV偍傛傃HTLV堚揱巕傪摫擖偟偨Tg儅僂僗傪嶌惢偟丄HIV-Tg偼僸僩偺寬忢僉儍儕儎乕偺儌僨儖偲偟偰丄HTLV堚揱巕摫擖儅僂僗偼娭愡儕僂儅僠偵傛偔帡偨枬惈娭愡墛偺儌僨儖偲偟偰桳梡偱偁傞偙偲偑傢偐偭偨丅杮峞偱偼HIV-Tg偵偮偄偰徯夘偡傞丅側偍丄HTLV-I偵偮偄偰偼暿偺徯夘婰帠乮娭愡儕僂儅僠偺敪徢婡峔丗幘姵儌僨儖偐傜偺峫嶡乯偑偁傞偺偱丄嶲徠偝傟偨偄丅
II丏僄僀僘
丂僄僀僘偼HIV姶愼偵傛傝丄CD4梲惈乮僿儖僷乕俿乯嵶朎偺尭彮偁傞偄偼婡擻晄慡偑堷偒婲偙偝傟丄偦偺寢壥柶塽椡偑掅壓偟丄庬乆偺擔榓尒姶愼偵傛偭偰巰偵帄傞昦婥偱偁傞丅僄僀僘姵幰偵尒傜傟傞kaposi擏庮傗僄僀僘擼徢偵傕HIV偑捈愙娭梌偡傞偲峫偊傜傟偰偄傞丅
丂HIV偼儗僩儘僂僀儖僗壢偺儗儞僠僂僀儖僗垷壢偵懏偡傞僂僀儖僗偱丄僾儔僗嵔RNA傪堚揱巕偲偟偰帩偮(2)丅姶愼屻丄媡揮幨峺慺偵傛傝俀杮嵔DNA偵曄姺偝傟丄廻庡愼怓懱偵慻傒崬傑傟丄堚揱巕敪尰傪峴偆丅堚揱巕偺椉抂偵偼LTR偲屇偽傟傞惂屼椞堟偑偁傝丄峔憿堚揱巕偲偟偰偼丄gag丄pol丄env側偳傪僐乕僪偟偰偄傞乮恾侾乯丅傑偨偙偺僂僀儖僗偼丄tat傗nef丄rev丄vpr側偳懡偔偺惂屼堚揱巕傪僐乕僪偟偰偄傞偙偲偑摿挜偱丄帺恎偺暋惢傪惂屼偡傞偩偗偱側偔丄廻庡偵傕暋嶨側塭嬁傪媦傏偡丅僂僀儖僗偺儗僙僾僞乕偲偟偰偼丄CD4暘巕偲摨帪偵CCR5傗CXCR4側偳偺働儌僇僀儞儗僙僾僞乕偑嫟摨偟偰婡擻偡傞偙偲偑抦傜傟偰偄傞丅儅僂僗偺CD4暘巕偼僸僩偺傕偺偲堎側偭偰偍傝丄HIV儗僙僾僞乕偲偟偰婡擻偟側偄丅偦傟偩偗偱側偔丄儅僂僗偲僸僩偱偼揮幨場巕側偳偵傕庬娫嵎偑偁傝丄偙偺偨傔丄HIV偼儅僂僗偱偼憹怋偱偒側偄偙偲偑傢偐偭偰偄傞丅
丂僄僀僘偼HIV偵姶愼偟偰偐傜捠忢俆乕侾侽擭偵傕偍傛傇挿偄愽暁婜傪宱偰敪徢偡傞丅嵟弶姶愼偟偨捈屻偵偼寣拞偵戝検偺僂僀儖僗偑専弌偝傟傞偑丄傗偑偰塼惈柶塽丄嵶朎惈柶塽偑惉棫偟偰寣拞偐傜偼僂僀儖僗偑傎偲傫偳徚幐偡傞丅偟偐偟偙偺帪揰偱傕儕儞僷愡偱偼僂僀儖僗偺嶻惗偑帩懕偟偰偄傞偙偲偑抦傜傟偰偍傝丄捠忢偺堄枴偱偺愽暁姶愼偱偼側偄丅偙偺娫丄僂僀儖僗偼偄偔偮偐偺撍慠曄堎傪婲偙偡偙偲偵傛傝丄柶塽傪摝傟傞偲嫟偵傛傝嫮偄撆惈傪妉摼偡傞丅堦曽僂僀儖僗偺憹怋偵傛偭偰CD4梲惈俿嵶朎偑攋夡偝傟傞偨傔偵丄彊乆偵俿嵶朎偑尭彮偟柶塽婡擻偑掅壓偡傞丅傗偑偰柶塽偲僂僀儖僗偲偺嬒峵偑攋傟傞偲媫寖偵寣拞僂僀儖僗検偑憹壛偟丄敪徢偵帄傞丅偙偺夁掱偵偍偄偰丄CD4梲惈嵶朎偺尭彮偲偲傕偵丄CD8梲惈嵶朎偺婡擻晄慡偑嵟嬤拲栚偝傟偰偍傝丄CD8嵶朎偑弌偡壗傜偐偺僂僀儖僗梷惂場巕偑暘斿偝傟側偔側傞壜擻惈傗丄Th1僞僀僾偐傜Th2僞僀僾傊偺僗僀僢僠偑峈僂僀儖僗妶惈偵拞怱揑側栶妱傪壥偨偟偰偄傞嵶朎惈柶塽偺掅壓偵娭梌偟偰偄傞偙偲偑峫偊傜傟偰偄傞丅傑偨丄徢忬偺埆壔偲僂僀儖僗検偼傛偔懳墳偡傞偙偲偑傢偐偭偰偍傝丄僂僀儖僗堚揱巕偺妶惈壔偼徢忬偺恑峴偵婎杮揑偵戝偒側栶妱傪壥偨偟偰偄傞偲峫偊傜傟傞丅
丂1995擭丄David Ho払偼僄僀僘姵幰偺側偐偱偼堦尒CD4梲惈俿嵶朎偺悢偑埨掕偟偰偄傞傛偆偵尒偊傞帪婜偱傕丄幚偼僂僀儖僗偑寖偟偔憹怋偟偰偍傝丄偙偺偨傔敿尭婜俀擔掱搙偲偄偆偒傢傔偰寖偟偄俿嵶朎偺turnover偑婲偙偭偰偄傞偙偲傪帵偡偲嫟偵丄 HIV偺曄堎昿搙偑嬌傔偰崅偔丄扨堦偺峈HIV栻偱偼偡偖偵懴惈曄堎僂僀儖僗偑偱偒偰偔傞偙偲傪帵偟偨(3)丅偙偺寢壥丄僄僀僘偺敪徢傪慾巭偡傞偵偼僂僀儖僗偺憹怋傪梷惂偡傞偙偲偑桳岠偱偁傝丄偟偐傕丄扨嵻偱偼側偔丄懡嵻傪摨帪偵搳梌偡傞偙偲偵傛偭偰懴惈僂僀儖僗偺弌尰傪梊杊偱偒傞偙偲偑傢偐偭偨丅偙偆偟偨抦尒偵婎偯偒丄懡嵻偵傛傞帯椕(HAART帯椕)偑峴傢傟傞傛偆側偭偨寢壥丄尰嵼偱偼僄僀僘偺敪徢傪偐側傝僐儞僩儘乕儖偱偒傞傛偆偵側偭偰偒偨丅偟偐偟丄巆擮側偙偲偵偙偺帯椕朄偱偼姶愼嵶朎傪姰慡偵嬱拃偡傞偙偲偼偱偒偢丄帯椕傪拞抐偡傞偲嵞傃僂僀儖僗偑憹怋偟巒傔傞偙偲偑抦傜傟偰偍傝丄傛傝崻杮揑側帯椕朄偺奐敪偑朷傑傟偰偄傞丅
III丏HIV堚揱巕摫擖Tg儅僂僗偺嶌惢
丂HIV-俬偺DNA僋儘乕儞NL4.3.2偐傜媡揮幨峺慺堚揱巕晹暘傪寚幐偝偣偨傕偺乮恾侾乯傪C3H/HeN儅僂僗偺庴惛棏偵儅僀僋儘僀儞僕僃僋僩偡傞帠偵傛傝嶌惢偟偨 乵C3H/HeN-TgN(HIV-儮pol)1607Jms乶(4)丅
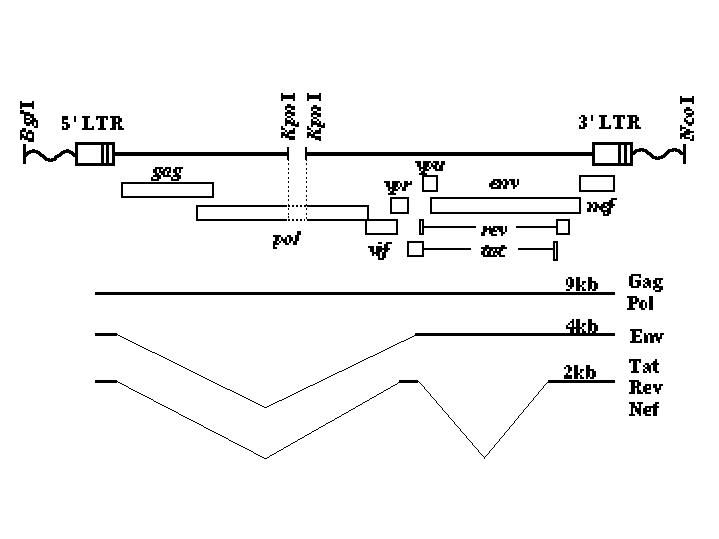
恾侾丏摫擖堚揱巕偺峔憿偲mRNA
丂HIV-1堚揱巕(NL4.3.2乯偺媡揮幨峺慺堚揱巕(Kpn I-Kpn I椞堟)傪寚懝偝偣偨傕偺傪摫擖偟偨丅
IV.HIV-1堚揱巕摫擖Tg儅僂僗偺摿挜
丂摫擖堚揱巕偼侾斣愼怓懱偺挿榬偺堦僇強偵栺侾侽僐僺乕偑憓擖偝傟偰偄偨丅偙傟傜偺儅僂僗偼俁僇寧楊崰偐傜敀撪忈傪敪徢偟巒傔丄12僇寧楊崰傑偱偵偼慡偰偺儅僂僗偑敪徢偟偨(4)丅枛徑寣拞偺CD4+/CD8+嵶朎斾偑10-40%掅壓偟偰偄偨偑丄棏敀傾儖僽儈儞偵懳偡傞柶塽墳摎擻偼惓忢偱偁偭偨丅摫擖堚揱巕偺敪尰偼娽偺儗儞僘嵶朎丄嬝擏側偳偱嫮偔尒傜傟丄旂晢傗擼丄銪憻丄嫻態側偳偱傕敪尰偑擣傔傜傟偨丅儊僢僙儞僕儍乕RNA偼僸僩姶愼嵶朎偱傒傜傟傞傕偺偲摨偠偔丄9kb丄4kb丄2kb偺俁庬偑擣傔傜傟偨丅僂僀儖僗峔惉抈敀幙傕惓忢偵崌惉偝傟偰偍傝丄寣拞偵偼惓忢側晜梀枾搙傪帩偮僂僀儖僗棻巕偑擣傔傜傟偨(5)丅
丂娽偺儗儞僘嵶朎偺拞偵偼戝検偺p24Gag抈敀幙偺拁愊偑擣傔傜傟偨偙偲偐傜丄偙偺偙偲偑敀撪忈偺尨場偱偁傞偲峫偊傜傟偨丅捠忢僄僀僘姵幰偵敀撪忈偑擣傔傜傟傞偙偲偼側偔丄Tg儅僂僗偱慡恎偺嵶朎偵堚揱巕偑慻傒崬傑傟偨偨傔偵傒傜傟傞尰徾偱偁傞丅堚揱巕偑慡恎偺嵶朎偵暘晍偟偰偄傞偵傕峉傢傜偢慻怐摿堎揑側堚揱巕敪尰偑尒傜傟偨偙偲偼丄HIV姶愼偺慻怐摿堎惈偑儗僙僾僞乕偩偗偱偼側偔丄嵶朎撪偺揮幨憰抲偵傛偭偰傕僐儞僩儘乕儖偝傟偰偄傞偙偲傪帵偟偰偄傞丅偙偲偵kaposi擏庮傗僄僀僘擼徢偺敪惗晹埵偱偁傞旂晢傗擼偱堚揱巕敪尰偑嫮偔擣傔傜傟偨偙偲偼嫽枴怺偄丅
丂儕儞僷慻怐偵墬偗傞摫擖堚揱巕偺敪尰偼惗棟忬懺偱偼旕忢偵掅偐偭偨偑丄嵶嬠偺儕億懡摐乮LPS乯傗僗乕僷乕峈尨乮SEB丗Staphylococcus Enterotoxin B)傪搳梌偡傞偲2-3擔屻偵敪尰偑30-40攞懀恑偝傟傞偙偲偑暘偐偭偨乮恾俀乯(5)丅LPS搳梌帪偵偼寣拞偵旕姶愼惈僂僀儖僗棻巕偑擣傔傜傟丄p24Gag抈敀幙偺寣拞擹搙偼僄僀僘姵幰偺寣拞擹搙偵旵揋偡傞丄400pg/ml偵傕払偟偨丅偙偺帪丄寣拞偵偼TNF-兛傗IL-1丄IL-6側偳偺墛徢惈僒僀僩僇僀儞偑崅儗儀儖専弌偝傟偨乮恾俁乯丅偦偙偱丄偙傟傜偺僒僀僩僇僀儞偑僂僀儖僗嶻惗偵媦傏偡塭嬁傪専摙偡傞偨傔偵丄僒僀僩僇僀儞寚懝儅僂僗偲偺妡偗崌傢偣傪峴偭偨丅偦偺寢壥丄TNF-兛丄偁傞偄偼IL-1兛/兝傪寚懝偝偣傞偲嫮偔HIV堚揱巕敪尰丄媦傃棻巕嶻惗偑梷惂偝傟傞偙偲偑傢偐偭偨乮恾係乯乮俆乯丅堦曽丄IL-1兛丄偁傞偄偼IL-1兝扨撈丄IL-6丄IFN-兞側偳偺寚懝偼堚揱巕敪尰偵塭嬁傪梌偊側偄偙偲偑傢偐偭偨丅偙偺寢壥丄HIV堚揱巕敪尰偵偼TNF-兛丄偍傛傃IL-1偑廳梫側栶妱傪壥偨偟偰偄傞偙偲偑傢偐偭偨丅
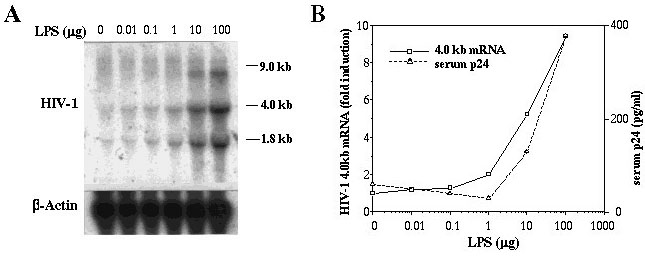
恾俀丏HIV堚揱巕偺LPS偵傛傞敪尰桿摫
HIV-Tg偵LPS傪暊峯撪搳梌偟偨偲偙傠丄梡検埶懚揑偵HIV堚揱巕偺敪尰桿摫偑傒傜傟偨丅俙丏銪憻RNA偺僲僓儞僴僀僽儕僟僀僛乕僔儑儞丄俛丏銪憻拞偺HIV摿堎揑mRNA検偲寣拞p24Gag抈敀幙検丅
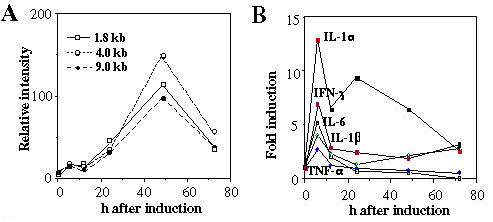
恾俁丏LPS搳梌屻偺HIVmRNA偺桿摫偲僒僀僩僇僀儞偺敪尰
HIV-Tg偵100mg偺LPS傪搳梌偟丄HIVmRNA偺敪尰(A)丄偍傛傃僒僀僩僇僀儞偺敪尰(B)傪宱帪揑偵應掕偟偨丅
恾係丏堚揱巕寚懝儅僂僗偵偍偗傞HIVmRNA 偺桿摫
偦傟偧傟偺僒僀僩僇僀儞寚懝HIV-Tg儅僂僗偵LPS傪搳梌偟丄HIVmRNA偺敪尰傪専摙偟偨丅TNF-兛丄偍傛傃IL-1兛/兝寚懝儅僂僗偱偼敪尰検偑掅壓偟偰偄偨丅
倁丏HIV-Tg偺幘姵儌僨儖偲偟偰偺桳梡惈
丂埲忋弎傋偨條偵丄HIV-Tg儅僂僗偵偍偄偰傕僸僩姶愼嵶朎偲摨偠偔惓忢側mRNA崌惉偑婲偙傝丄偐偮丄偦偺mRNA偐傜惓忢側僂僀儖僗棻巕抈敀幙偑嶌傜傟傞偙偲偑傢偐偭偨丅偟偐偟側偑傜丄媡揮幨峺慺傪寚懝偝偣偰偄傞偨傔丄摉慠偺偙偲偲偟偰偙傟傜偺棻巕偵姶愼惈偼擣傔傜傟側偐偭偨乮俆乯丅偙偺儅僂僗偱偼捠忢儕儞僷慻怐偵偍偗傞堚揱巕敪尰偺儗儀儖偼掅偔丄LPS側偳偱妶惈壔偡傞偙偲偵傛偭偰丄嫮偔敪尰偑桿摫偝傟傞丅廬偭偰丄偙偺Tg儅僂僗偼僸僩偺寬忢姶愼幰偺儌僨儖偲峫偊傞偙偲偑偱偒傞丅埲壓偺條側棙梡偑峫偊傜傟傞丅
侾乯峈HIV栻偺僗僋儕乕僯儞僌丅
丂偙傟傑偱丄彫摦暔傪巊偭偨僗僋儕乕僯儞僌宯偼側偔丄杮Tg儌僨儖偼嬌傔偰婱廳偱偁傞丅嵶朎儗儀儖偩偗偱偼撆惈傗戙幱偺栤戣偑偁傞偨傔丄摦暔屄懱傪巊偭偨専摙偑昁梫偱偁傝丄僒儖側偳傪巊偭偰峴偆専摙偺慜抜奒偲偟偰桳梡偱偁傞丅
俀乯HIV堚揱巕偺妶惈壔偺儊僇僯僘儉偺夝愅丅
丂摦暔屄懱傪巊偭偰HIV偺妶惈壔偑傒傜傟傞宯偼楈挿椶偺懠偵偼側偄丅摿偵丄儅僂僗偺応崌丄堚揱攚宨丄旝惗暔墭愼傪尩枾偵惂屼偱偒傞傎偐丄懠偺僩儔儞僗僕僃僯僢僋儅僂僗傗僲僢僋傾僂僩儅僂僗偲妡偗崌傢偣傞側偳偺丄僒儖偱偼偱偒側偄傾僾儘乕僠偑壜擻偱偁傞丅
俁乯HIV偺昦尨惈偺専摙丅
丂崅儗儀儖偺僂僀儖僗嶻惗偑傒傜傟傞偙偲偐傜丄廻庡偵懳偡傞撆惈傪専摙偡傞偨傔偵桳梡偱偁傞丅幚嵺丄偙傟傜偺儅僂僗偼惗棟忬懺偱CD4梲惈嵶朎偑10?40亾尭彮偟偰偍傝丄姶愼姵幰偲摨偠條側偙偲偑婲偒偰偄傞傕偺偲峫偊傜傟傞丅廬偭偰丄偙傟傜偺儅僂僗傪巊偆偙偲偵傛傝丄俿嵶朎尭彮偺儊僇僯僘儉偺夝愅傗丄姶愼俿嵶朎摦懺偺夝愅丄偝傜偵偼姶愼俿嵶朎攔彍朄偺専摙側偳丄懡偔偺壽戣傪専摙偡傞偙偲偑偱偒傞傕偺偲峫偊偰偄傞丅
丂偲偙傠偱丄杮儌僨儖偺堚揱巕敪尰検偼姶愼僸僩嵶朎偵妑傋丄悢侾侽暘偺侾掱搙偟偐側偄丅偙傟偼揮幨場巕偺堦偮偱偁傞丄Cyclin T1偑僸僩偲儅僂僗偱堎側偭偰偄傞偨傔偱偁傞壜擻惈偑峫偊傜傟偨丅偦偙偱丄Cyclin T1堚揱巕傪摫擖偟偨Tg儅僂僗傪嶌惢偟偨偲偙傠丄僂僀儖僗偺敪尰検偑100~1000攞傕槾恑偡傞偙偲偑傢偐偭偨乮6乯丅偙偺僟僽儖Tg儅僂僗傪梡偄傞偙偲偵傛傝丄傛傝徻嵶側昦尨惈偺夝愅偑壜擻偵側傞傕偺偲婜懸偟偰偄傞丅
暥專
1. Sugamura, K., and Hinuma, Y.: Human retroviruses:HTLV-I and HTLV-II. In"The Retroviridae", vol. 2, ed. J. A. Levy, Plenum Press, New York, 1993, 399-435.
2. Levy, J. A.: Pathogenesis of human immunodeficiency virus infection. Microbiol. Reviews 1993, 57:183-289.
3.Ho, D. D., Neumann, A. U., Perelson, A. S. et al.: Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature 1995, 373:123-126.
4. Iwakura, Y., Shioda, T., Tosu, M., et al.: Human immunodeficiency virus type 1 induces cataract in transgenic mice. AIDS, 1992, 6:1069-1075.
5. Tanaka, J., Ozaki, H., Yasuda, J., et al.: Lipopolysaccharide-induced HIV-I expression in transgenic mice is mediated by tumor necrosis factor-a and interleukin-1, but not by interferon-g nor interleukin-6. AIDS, 2000, 14:1299-1307.
6.埨揷擇楴丄揷拞弮丄娾憅梞堦榊丄戞48夞擔杮僂僀儖僗妛夛彺榐廤丄2000丄p.259丅