研究成果 注目の論文
環境記憶統合 “注目の論文” No. 16 植物に乾燥・高温耐性を付与するDREB2Aが活性化する仕組みを解明
-
- 論文名
- BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis
- 著者名(下線は環境記憶統合メンバー)
- #Kyoko Morimoto, #Naohiko Ohama, Satoshi Kidokoro, Junya Mizoi, Fuminori Takahashi, Daisuke Todaka, Junro Mogami, Hikaru Sato, Feng Qin, June-Sik Kim, Yoichiro Fukao, Masayuki Fujiwara, Kazuo Shinozaki, and *Kazuko Yamaguchi-Shinozaki
#These authors contributed equally to this work. - 雑誌名等
- Proceedings of the National Academy of Sciences of the United States of America, 113 (32), 8969-8974 (2017), doi:10.1073/pnas.1704189114
http://www.pnas.org/content/114/40/E8528.full - 解説
-
地球レベルの環境劣化が進み、異常気象による農業被害が深刻化しています。さらに地球温暖化の影響によって、急激な温度変化や干ばつなどが頻発するようになると予測されています。このため、環境ストレスに高い耐性を持つ作物の育種が重要となっています。植物は、環境ストレスを受けると、さまざまな遺伝子の発現を変化させることにより耐性を獲得します。DREB2Aは、高温や乾燥の複合ストレスに対する植物の耐性獲得機構において中心的な役割を果たす転写因子で、植物が乾燥と高温の両方のストレスに対する耐性を獲得するために機能する多数の遺伝子群の働きを活性化させることができます。このため、環境ストレス耐性作物の分子育種における有望な因子と考えられています。しかし、DREB2Aをそのまま植物中で強く発現させても働きを示しません。私たちの研究グループは10年前、DREB2Aの構造からNegative regulatory domain (NRD)と名付けた30アミノ酸の配列を取り除くと活性型(DREB2A CA)に変換できることを発見しました。さらに、DREB2A CAを植物中で高発現すると、多数の標的遺伝子の発現が強く誘導されて、乾燥と高温ストレスに対する耐性が向上することも示しました。しかし、なぜNRD配列を除くとDREB2Aは活性型に変換されるのか、その分子メカニズムは長い間不明のままでした。
今回、私たちのグループは、シロイヌナズナを用いてDREB2Aのタンパク質複合体を精製し、質量分析装置を用いることでDREB2Aと相互作用するタンパク質を解析しました。その結果、DREB2A相互作用タンパク質として、CUL3 E3リガーゼの基質アダプターであるBPMを同定しました。BPMはDREB2Aと核内で相互作用し、この時、NRD配列がBPMの認識・結合部位として働くことを見出しました。また、BPMの働きを抑えた植物を用いた実験から、BPMはNRD依存的にDREB2Aの分解を促進することが示されました。これらの結果から、NRDがDREB2Aを不安定化する分子メカニズムは、BPM-CUL3のリクルートであると考えられます(図1)。
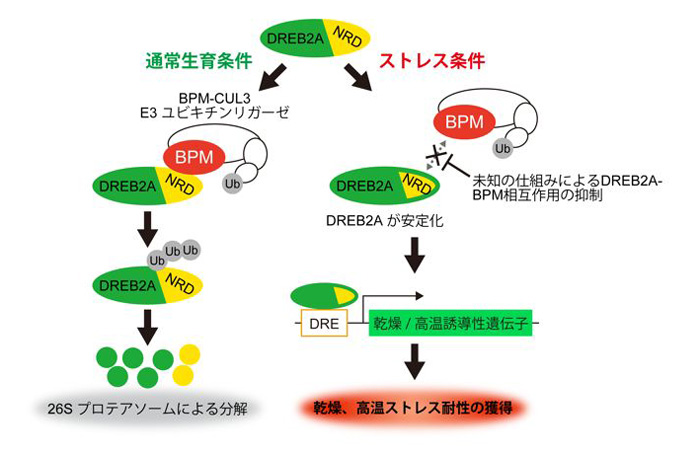
図1 植物の干ばつと高温の両方の耐性獲得に働く転写因子DREB2Aの構造中のNRDは、タンパク質の分解系であるBPM-CUL3 E3リガーゼが認識して結合する配列である。通常生育条件下では、その結合によりDREB2Aはユビキチン化されプロテアソーム系に運ばれ分解される。乾燥・高温ストレス時にはリン酸化などによりNRDが修飾されたり、または未知のタンパク質によってNRDがマスクされたりして、BPM-CUL3 E3リガーゼによる分解系から逃れることで活性化する。活性化したDREB2Aは耐性遺伝子の発現を誘導し乾燥や高温耐性が獲得される。
BPMの働きを抑えた植物を高温または乾燥ストレス処理すると、DREB2Aタンパク質が顕著に高蓄積しました。その結果、DREB2Aの標的遺伝子群が強く発現誘導され、高温耐性が向上しました(図2)。
このため、BPM-CUL3は通常生育条件、ストレス条件を通してDREB2Aのタンパク質レベルを厳密に制御する仕組みであると考えられます。
図2 新たに同定された、CUL3 E3リガーゼの基質アダプターであるBPMの働きを抑えた植物(amiBPM)では、高温耐性の向上が示された。
高温などのストレス時の植物中では、リン酸化などのNRDの修飾や未知のタンパク質によるNRDのマスクにより、BPM-CUL3 E3リガーゼがDREB2Aを認識できなくなると推測されます(図1)。本研究では、質量分析装置を用いてDREB2Aタンパク複合体を分析することで、BPM-CUL3 E3リガーゼとDREB2Aの関係を明らかにすることに成功しました。さらに、様々な転写因子においてもNRDに類似した配列が見出されることから、代謝回転が早い転写因子の分解制御においてBPM-CUL3 E3リガーゼが重要な役割を果たしていることが考えられました。
以上の結果より、これまで10年以上の長い間不明のままであったDREB2A CAの活性化の機構が明らかになりました。今回、DREB2Aの相互作用因子として単離されたBPM-CUL3 E3リガーゼは、DREB2Aタンパク質をユビキチン化してプロテアソーム分解系に運ぶシステムであり、NRD配列を特異的に認識して結合しDREB2Aを分解系に運びます。NRDを持たないDREB2A CAは、分解系から逃れ安定化することで活性化することが示されました。DREB2Aは多くの耐性遺伝子群を制御して乾燥と高温の両方のストレス耐性を向上しますが、成長には負の影響を示すことが明らかにされています。そのため、不必要な時はBPM-CUL3 E3リガーゼによりユビキチン化してタンパク質の分解系に運んでいると考えられました。これらのシステムを効率よく制御することで、高温や干ばつに対する耐性を向上させた作物の開発への応用が期待されます。