研究成果 注目の論文
環境記憶統合 “注目の論文” No. 19 窒素栄養に応答して根粒形成を抑制する鍵転写因子NRSYM1の発見
-
- 論文名
- A NIN-LIKE PROTEIN mediates nitrate-induced control of root nodule symbiosis in Lotus japonicus
- 著者名(下線は環境記憶統合メンバー)
- Hanna Nishida, Sachiko Tanaka, Yoshihiro Handa, Yuki Sakamoto, Sachihiro Matsunaga, Shigeyuki Betsuyaku, Kenji Miura, Takashi Soyano, Masayoshi Kawaguchi, and *Takuya Suzaki
- 雑誌名等
- Nature Communications, 9, 499 (2018) (14 pages), doi: 10.1038/s41467-018-02831-x
https://www.nature.com/articles/s41467-018-02831-x - 解説
-
植物が生きる上で必須の栄養素である窒素は大気中の約80%を占めますが、多くの植物はその窒素を直接利用することができないため、土壌中に存在する窒素栄養(主に硝酸やアンモニア)を利用して生きています。マメ科植物は、根に根粒と呼ばれるこぶ状の器官を形成し、その中に共生する根粒菌が大気中の窒素を植物が利用できる形に変換してくれる(窒素固定)ので、土壌中の窒素栄養が乏しい環境でも生育することが可能です。しかし、根粒菌との共生を行うためには、窒素固定反応などのエネルギー源として光合成産物を根粒菌へ供給しなければなりません。そこで、宿主植物は共生を沢山おこなって光合成産物を消費しすぎることで成長に悪影響を及ぼさないように、自身の根につくる根粒の数を適切に保つ仕組みをそなえています。また、土壌中に窒素栄養が豊富に存在する場合には、そちらを優先的に利用することができるので、エネルギーのかかる根粒菌との共生をやめて光合成産物の消費を抑えています。この、土壌中の窒素栄養に応答した根粒共生の抑制は古くから知られた現象であるものの、その制御機構についての理解はあまり進んでいませんでした。
本研究では、マメ科のモデル植物ミヤコグサを用いて、窒素栄養(硝酸)が豊富に存在する土壌でも根粒を形成するような突然変異体のスクリーニングを行い、nitrate unresponsive symbiosis 1 (nrsym1)と名付けた新しい変異体を単離しました。nrsym1変異体は窒素栄養を含まない土壌では野生型と同様の根粒共生を示します。高濃度の硝酸を添加した場合、野生型の植物は根粒を作らなくなりますが、nrsym1変異体は窒素栄養を含まない条件とほとんど変わらない成熟した根粒を形成しました(図1)。したがって、NRSYM1遺伝子は高濃度の硝酸に応答して引き起こされる根粒共生の抑制を制御する因子であることがわかりました。nrsym1変異体の原因遺伝子を特定したところ、NIN-LIKE PROTEIN (NLP)と呼ばれるタイプの転写因子をコードしていることが明らかになりました。シロイヌナズナなどの研究から、NLPは硝酸に応答して様々な遺伝子の発現を制御する植物の硝酸応答の中心的な役割を担う因子であることが示唆されています。
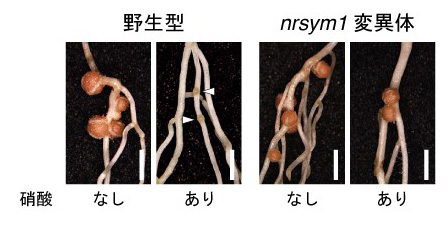
図1 野生型植物とnrsym1変異体の根粒共生。矢じりは未熟な根粒を示す。スケールバー:2 mm
これまでの研究から、宿主植物が備えている根粒の数を適切に保つ仕組みは、Autoregulation of nodulation (AON)と呼ばれる根と地上部を介した全身的なシグナル伝達によって制御されていることが明らかにされています。そしてAONが窒素栄養に応答した根粒共生の抑制の制御にも関わっている可能性が示唆されていました。野生型植物では、AONにおいて根粒菌の感染を根から地上部へ伝えるためのシグナルとして働くCLEペプチドの一つであるCLE-RS2遺伝子の発現が根粒菌の感染だけでなく硝酸によっても強く誘導されます。一方でnrsym1変異体では硝酸添加によるCLE-RS2遺伝子の発現誘導が阻害されたため、NRSYM1転写因子が硝酸に応答したCLE-RS2遺伝子の発現を制御している可能性が考えられました。そこで、CLE-RS2遺伝子がNRSYM1の直接の標的遺伝子であるかを検証したところ、硝酸添加時にNRSYM1がCLE-RS2のプロモーター上に結合することが確認されました。これらの結果から、NRSYM1転写因子は硝酸に応答してCLE-RS2遺伝子の発現を直接誘導し、AONと同様の機構を用いて全身的に根粒の数を制御していることが明らかになりました(図2)。
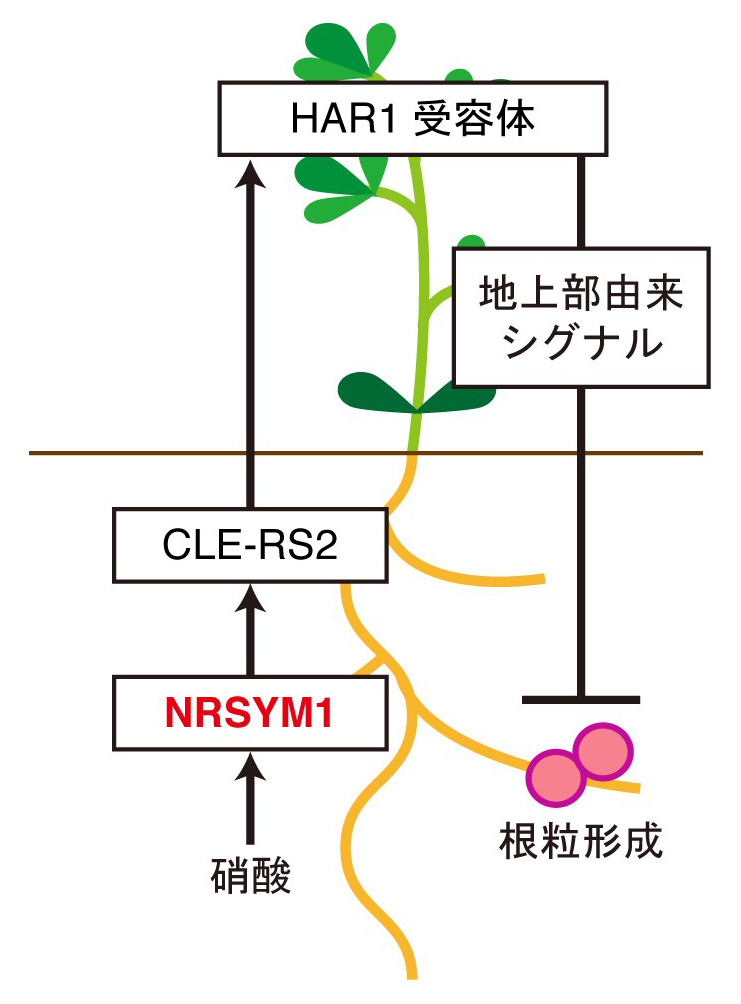
図2 NRSYM1転写因子によるAONを介した全身的な根粒数の制御機構のモデル図
硝酸に応答したNRSYM1は根においてCLE-RS2遺伝子の発現を直接誘導する。地上部へと移動したCLE-RS2がHAR1受容体によって受容された後、地上部由来シグナルが生産されて根へと移動し根粒形成を抑制する。また、NRSYM1タンパク質の細胞内局在を調べてみると、NRSYM1タンパク質は硝酸添加時には核に局在し、無窒素の条件では核への局在がほとんど見られなくなることがわかりました(図3)。したがって無窒素条件では核の外にいるNRSYM1が、硝酸に応答して核に移行し、標的遺伝子の発現を誘導することで、硝酸に応答した根粒共生の抑制を引き起こす可能性が示唆されました。
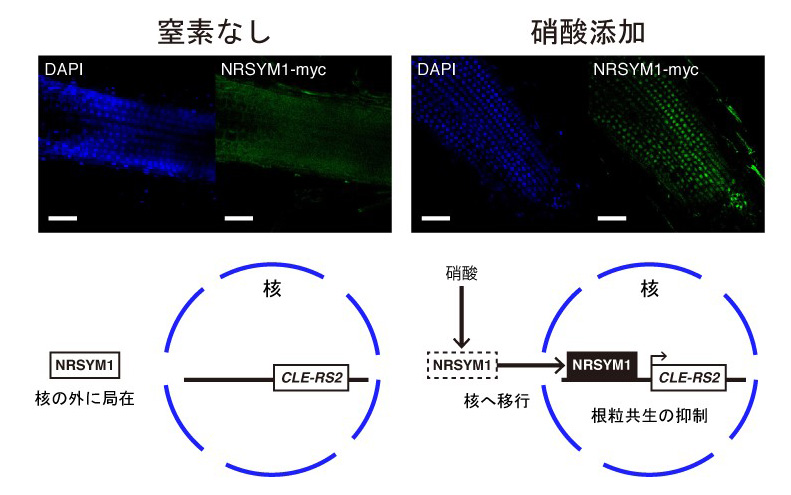
図3 NRSYM1タンパク質の細胞内局在と遺伝子発現制御モデル
核の位置をDAPI染色(青色)、NRSYM1タンパクの局在をMyc抗体を用いた抗体免疫染色(緑色)によってそれぞれ可視化している。スケールバー:50 μm
無窒素の条件では核の外に局在しているNRSYM1は、硝酸に応答して核へと移行し、標的遺伝子のプロモーターに結合して発現を誘導することにより根粒共生を抑制する。本研究によって、NRSYM1転写因子が硝酸に応答した根粒共生制御の中心的な働きを担うことが明らかになりました。NRSYM1は様々な下流因子を使い分けることで根粒共生を多面的に制御していると予想されます。今後はCLE-RS2遺伝子以外のNRSYM1標的遺伝子を同定することで、硝酸に応答した根粒共生の制御機構の全容解明に迫っていきたいと考えています。また、農業の現場においては作物を育てるために窒素肥料が使用されていますが、過剰量の窒素肥料施肥による環境汚染などが問題視されています。そこで根粒共生を利用した自然に優しい窒素源の獲得が注目されていますが、共生による窒素固定のみで作物が十分に生育するための窒素源を確保することは困難です。窒素栄養に応答した根粒共生の抑制の仕組みを理解することは、窒素肥料と根粒共生による窒素源の獲得の両立のために重要であり、NRSYM1の研究から得られた知見は持続可能な農業の実現に貢献できる可能性が期待できます。